 |
| Cerci of Opisthocosmia cervipyga (Forficulidae: Opisthocosmiinae); photographed by Fabian Haas (©BMNH) |
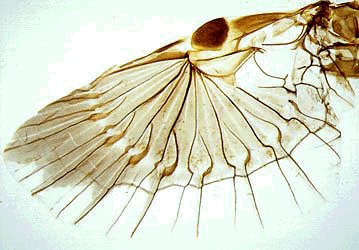 |
| Outstretched hind wing of Allodahlia scabriscula (Forficulidae) |
 |
| Hemimerus vosseleri (female) |
 |
| Arixenia esau (female) |
Both of these families have adapted to an epizoic lifestyle by developing live birth (vivipary)—with embryos each developing at the termini of ovarian follicles, nourished by a thickened epithelium dubbed a "pseudoplacenta" (Hagan, 1951). Gestation in Arixenia esau is additionally complicated by the movement of older fetuses into their mother's oviduct, which is modified into an ad hoc uterus (Tworzydlo et al., 2013). It probably goes without saying that other earwigs conventionally lay eggs, except for the unexplained aberration that is Chaetospania borneensis (Spongiphoridae) (Kočárek, 2009).
 |
| An Apachyus sp. (Apachyidae); purportedly one of the closest relatives of the Hemimeridae (Engel & Haas, 2007) |
Hence, for further illumination of earwig evolution we must draw on data derived from the field of paleontology, despite the fact that the dermapteran fossil record is far from copious (Wappler et al., 2005). Nevertheless, it includes not only representatives of the extant superfamilies Pygidicranoidea—putatively dating as far back as the late Jurassic (Shcherbakov, 2002) or no later than the mid-Cretaceous (Engel & Grimaldi, 2004)—and Forficuloidea (Wappler et al., 2005), as well as members of the similarly living families Labiduridae (Zhao et al., 2010) and Diplatyidae (Engel, 2011); but also two wholly prehistoric suborders: the Archidermaptera (Jurassic of Eurasia; Whalley, 1985) and Eodermaptera (later Jurassic of Kazakhstan and Manchuria, perhaps extending to as late as the Cretaceous; Zhang, 1994; Zhao et al., 2010).
The latter suborder is evidently akin to the Neodermaptera, as deduced from their shared lack of annulation in the cerci (Haas & Kukalová-Peck, 2001) and 3-3-3 tarsal formula‡, but differ in their retention of ocelli (Zhao et al., 2010). In their turn, the Archidermaptera (which include five families; Grimaldi & Engel, 2005) are distinguished from the remainder of the order by the presence of venation in their tegmina and primitively segmented cerci: although it should be noted here that nymphs of Karschiellidae and Diplatyidae (Neodermaptera) do also retain like segmentation (Burr, 1911; Shimizu & Machida, 2011b), hinting at already apparent paraphyly among these ancient earwigs (Willmann, 1990).
 |
| Mother F. auricularia from Washington protecting her eggs from the photographer (Lynette Schimming) |
But enough of the evolutionary biography of earwigs as a taxon; what of their lineage? No self-respecting overview of the scientific opinion on this subject could omit mention of the diverse order Protelytroptera, which are temporally restricted to the Permian Period and crop up throughout what was then the supercontinent of Pangaea (Kukalová, 1966). Formerly, the classification of the Umenocoleidae herein (Carpenter, 1992) extended protelytropterans' range beyond the massive Permo-Triassic extinction (Arnett, 2000): but umenocoleids have since been unmasked as bizarre cockroaches (Vršanský et al., 2002). The most salient protelytropteran trait would be their sclerotized forewings, which paleoentomologists even go so far as to term "elytra" in likeness of the analogous (but not homologous) features for which beetles are known: indeed, several protelytropterans were once upon a time mistaken for some form of quasi-beetle (Tillyard, 1924; Laurentiaux, 1953). Wing structures in the order are suggestive of an affinity with earwigs, and it is as such held that the Dermaptera descend from within the Protelytroptera (Haas & Kukalová-Peck, 2001): but this theory remains unconfirmed.
Other than their inclusion in the clade Polyneoptera, we can say little and know less of the broader relations of this ordinal duo: proposals vaguely lean towards distant kinship with the Notoptera, Dictyoptera (mantids/cockroaches/termites), Zoraptera, Plecoptera (stoneflies), or selections from the above in one permutation or another (Boudreaux, 1979; Haas & Kukalová-Peck, 2001; Jarvis et al., 2005; Terry & Whiting, 2005; Wheeler et al., 2001; Simon et al., 2010). In the end, the jury is still out as to the exact origin of earwigs.
I could have told you so right out of the gate; but where's the fun in that?
*As this would refer to colugos, which belong in a wholly different superphylum.
†The trio of ommatidia situated between the compound eyes in the insect cephalic groundplan.
‡Which is to say that all of their tarsi consist of five segments.
___________________________________________________________
Burr, M. (1911). Dermaptera. Genera Insectorum, 122, 1-112.
Boudreaux, H. B. (1979). Arthropod Phylogeny with Special Reference to Insects. John Wiley & Sons: New York.
Carpenter, F. M. (1992). Treatise on Invertebrate Paleontology (vol. 3, pt. R). Boulder: Geological Society of America.
Engel, M. S. (2003). The earwigs of Kansas, with a key to genera north of Mexico [electronic version]. Transactions of the Kansas Academy of Science, 106(3/4), 115-123. Retrieved 2/15/14 from http://www.jstor.org/stable/3628391?seq=1
Engel, M. S. (2011). New earwigs in mid-Cretaceous amber from Myanmar (Dermaptera, Neodermaptera). ZooKeys, 130, 137-152. Retrieved 2/15/14 from http://www.pensoft.net/J_FILES/1/articles/1293/1293-G-3-layout.pdf
Engel, M. S. and Grimaldi, D. (2004). A primitive earwig in Cretaceous amber from Myanmar (Dermaptera: Pygidicranidae). Journal of Paleontology, 78(5), 1018-1023.
Engel, M. S. and Haas, F. (2007). Family-group names for earwigs (Dermaptera) [electronic version]. American Museum Novitates, 3567, 1-20. Retrieved 2/15/14 from http://digitallibrary.amnh.org/dspace/bitstream/handle/2246/5858//v3/dspace/updateIngest/pdfs/N3567.pdf?sequence=1
Giles, E. T. (1963). The comparative external morphology and affinities of the Dermaptera. Transactions of the Royal Entomological Society of London, 115, 95-164.
Grimaldi, D. and Engel, M. S. (2005). Evolution of the Insects. Cambridge: Cambridge University Press.
Haas, F.; Gorb, S. N.; and Wootton, R. J. (2000). Elastic joints in dermapteran hind wings. Materials and wing folding. Arthropod Structure and Development, 29, 137-146.
Haas, F. and Kukalová-Peck, J. (2001). Dermaptera hindwing structure and folding: new evidence for familial and superordinal relationships within Neoptera (Insecta). European Journal of Entomology, 98, 445-509.
Haas, F. and Klass, K. D. (2003). The basal phylogenetic relationships in the Dermaptera. In Klass, K. D. (ed.): Proceedings of the 1st Dresden meeting on insect phylogeny: "Phylogenetic Relationships within the Insect Orders" (Dresden, September 19-21, 2003). Entomologische Abhandlungen, 61, 132-133.
Hagan, H. R. (1951). Embryology of Viviparous Insects. New York City: Ronald Press.
Hand, W. D. (1976). From idea to word: folk beliefs and customs underlying folk speech [electronic version]. American Speech, 48(1/2), 67-76. Retrieved 1/25/14 from http://www.jstor.org/stable/3087894
Jarvis, K. J.; Haas, F.; and Whiting, M. F. (2005). Phylogeny of earwigs (Insecta: Dermaptera) based on molecular and morphological evidence: reconsidering the classification of Dermaptera. Systematic Entomology, 30, 442-453.
Kočárek, P. (2009). A case of viviparity in a tropical non-parasitizing earwig (Dermaptera: Spongiphoridae). Tropical Zoology, 22, 237-241.
Kočárek, P.; Vaclav, J.; and Hulva, P. (2013). When the body hides the ancestry: phylogeny of morphologically modified epizoic earwigs based on molecular evidence. PLoS One. Retrieved 2/15/14 from http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0066900
Kukalová, J. (1966). Protelytroptera From the Upper Permian of Australia, With a Discussion of the Protocoleoptera and Paracoleoptera [electronic version]. Psyche, 73(2), 89-111. Retrieved 2/20/14 from http://www.hindawi.com/journals/psyche/1966/093549/abs/
Laurentiaux, D. (1953). Insects. In Piveteau, J. (ed.): Traité de Paléontologie, 3, 397-527.
Medway, L. (1958). On the habit of Arixenia esau Jordan (Dermaptera). Proceedings of the Royal Entomological Society of London (A), 33, 191-195.
Nakata, S. and Maa, T. C. (1974). A review of the parasitic earwigs (Dermaptera, Arixeniina; Hemimerina). Pacific Insects, 16, 307-374.
Popham, E. J. (1961). On the systematic position of Hemimerus Walker—a case for ordinal status. Proceedings of the Royal Entomological Society of London, Series B; 30, 19-25.
Popham, E. J. (1985). The mutual affinities of the major earwig taxa (Insecta: Dermaptera). Zeitschrift fur Zoologische Systematik und Evolutionforschung, 23, 199-214.
Rehn, J. A. G. and Rehn, J. W. H. (1935). A study of the genus Hemimerus (Dermaptera, Hemimerina, Hemimeridae) [electronic version]. Proceedings of the Academy of Natural Sciences of Philadelphia, 87, 457-508. Retrieved 2/15/14 from http://www.jstor.org/stable/4064223
Rentz, D. C. F. and Kevan, D. K. M. (1991). Dermaptera. In Naumann, D. T. (ed.) (pp. 360-368): Insects of Australia. Melbourne: CSIRO.
Sakai, S. (1982). A new proposed classification of the Dermaptera with special reference to the checklist of the Dermaptera of the world. Bulletin of Daito Bunka University, 20, 1-108.
Sharp, D. (1892). Concerning Hemimerus talpoides Walker. Entomological Monthly Magazine, 28, 212-213.
Shcherbakov, D. E. (2002). Order Forficulida Latreille, 1810. The earwigs and protelytropterans (=Dermaptera DeGeer, 1773+Protelytroptera Tillyard, 1931). In Rasnitsyn, A. P. and Quicke, D. L. J. (eds.) (pp. 288-291): History of Insects. Dordrecht: Kluwer Academic Publishers.
Shimizu, S. and Machida, R. (2011a). Notes on mating and oviposition of a a primitive representative of the higher Forficulina, Apachyus chartaceus (de Haan) (Insecta: Dermaptera: Apachyidae). Arthropod Systematics & Phylogeny, 69(2), 75-81. Retrieved 2/19/14 from http://www.senckenberg.de/files/content/forschung/publikationen/arthropodsystematics/asp_69_2/asp_69_2_shimizu_75-81.pdf
Shimizu, S. and Machida, R. (2011b). Reproductive biology and postembryonic development in the basal earwig Diplatys flavicollis (Shiraki) (Insecta: Dermaptera: Diplatyidae). Arthropod Systematics & Phylogeny, 69(2), 83-97. Retrieved 2/19/14 from http://www.senckenberg.de/files/content/forschung/publikationen/arthropodsystematics/asp_69_2/asp_69_2_shimizu_83-97.pdf
Simon, S.; Schierwater, B.; and Hadrys, H. (2010). On the value of elongation factor-1α for reconstructing pterygote insect phylogeny. Molecular Phylogenetics and Evolution, 54, 651-656.
Steinmann, H. (1986). Dermaptera: Catadermaptera I. Wermuth, H. and Mühn, E. (eds.): Das Tierreich 102. Berlin: de Gruyter.
Terry, M. D. and Whiting, M. F. (2005). Mantophasmatodea and phylogeny of the lower neopterous insects. Cladistics, 21, 240-257.
Tillyard, R. J. (1924). Upper Permian Coleoptera and a new order from the Belmont Beds, New South Wales. Proceedings of the Linnean Society of New South Wales, 42(4), 720-756.
Tworzydlo, W.; Kisiel, E.; and Bilinski, S. M. (2013). Embyros of the viviparous dermapteran, Arixenia esau develop sequentially in two compartments: terminal ovarian follicles and the uterus. PLoS One. Retrieved 2/15/14 from http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0064087
Vishnyakova, V. N. (1980). Earwigs from the Upper Jurassic of the Karatau range. Paleontological Journal, 1, 78-95.
Vršanský, P.; Vishnyakova, V. N.; and Rasnitsyn, A. P. (2002). Order Blattida Latreille, 1810. In Rasnitsyn, A. P. and Quicke, D. L. J. (eds.) (pp. 263-270): History of Insects. Dordrecht: Kluwer Academic Publishers.
Walker, F. (1871). Catalogue of the Specimens of Dermaptera Saltatoria in the Collection of the British Museum (pt. 5). London.
Wappler, T.; Engel, M. S.; and Haas, F. (2005). The earwigs (Dermaptera: Forficulidae) from the middle Eocene Eckfeld maar, Germany. Polskie Pismo Entomologiczne, 74(3), 227-250.
Whalley, P. E. S. (1985). The systematics and palaeogeography of the Lower Jurassic insects of Dorset, England. Bulletin of the British Museum of Natural History (Geology Series), 39, 107-189.
Wheeler, W. C.; Whiting, M.; Wheeler, Q. D.; and Carpenter, J. M. (2001). The phylogeny of the extant hexapod orders. Cladistics, 17, 113-169.
Willmann, R. (1990). Die Bedeutung paläontologischer Daten für die zoologische Systematik. Verhandlungen der Deutschen Zoologischen Gesellschaft, 83, 277-289.
Zhang, J. F. (1994). Discovery of primitive fossil earwigs (Insecta) from Late Jurassic of Laiyang, Shandong and its significance. Acta Palaeontologica Sinica, 33, 229-245.
Zhao, J.; Zhao, Y.; Shih, C.; Ren, D.; and Wang, Y. (2010). Transitional fossil earwigs—a missing link in Dermaptera evolution. BMC Evolutionary Biology, 10(344). Retrieved 2/15/14 from http://www.biomedcentral.com/content/pdf/1471-2148-10-344.pdf