While you contemplate that title, allow me to explain the subject of this installment: previously, I discussed the multitudinous fauna that inhabit the abodes of social insects, and sometimes reap the benefits of membership in said insect societies by impersonating these insects. But there is another aspect of societal symbioses among the Insecta; in many cases, both symbionts are social (or at least descend from eusocial ancestors), producing fascinating compound societies in the process. Originally, the phrase "social parasitism" referred only to these cases (Forel, 1898).
In its most basic form, inter-specific relation is simply colonial cohabitation (parabiosis). One exemplar of this habit would be the termites of the genus Inquilinitermes, which appear to be exclusive inquilines of Constrictotermes cyphergaster (although there is a single record of an Inquilinitermes colony residing without association with any other termite; Apolinário & Martius, 2004): their colonies (less populous than those of the host) occupy limited portions of Constrictotermes termitaria, and Inquilinitermes reside in galleries distinct from their hosts' (Cunha et al., 2003): these contain an excess of decomposed organic materials, upon which the inquilines probably subsist (Mathews, 1977).
 |
| 3 nests of C. cyphergaster from São João do Cariri (Brazil) (Bezerra-Gusmão et al., 2011) |
Inquiline termites (of which there are a number besides the genus previously mentioned) present an unusual ecological interface with their fellow isopteran hosts, in that both participants in the symbiosis possess the same diet (contrary to the synecthrans described in Pt. 1); isotopic analysis of several builder-inquiline pairs shows that there is careful dietary segregation: to be expected, given the colonies' spatial partitioning (Florencia et al., 2013). Not directly interacting with the architects of their abodes, many termite inquilines are better termed "termitariophiles" (Berg, 1900): an epithet that could also be applied to the African ant Carebara vidua, belonging to a speciose pantropical (Brown, 2000) genus of tiny, poorly-known myrmicines.
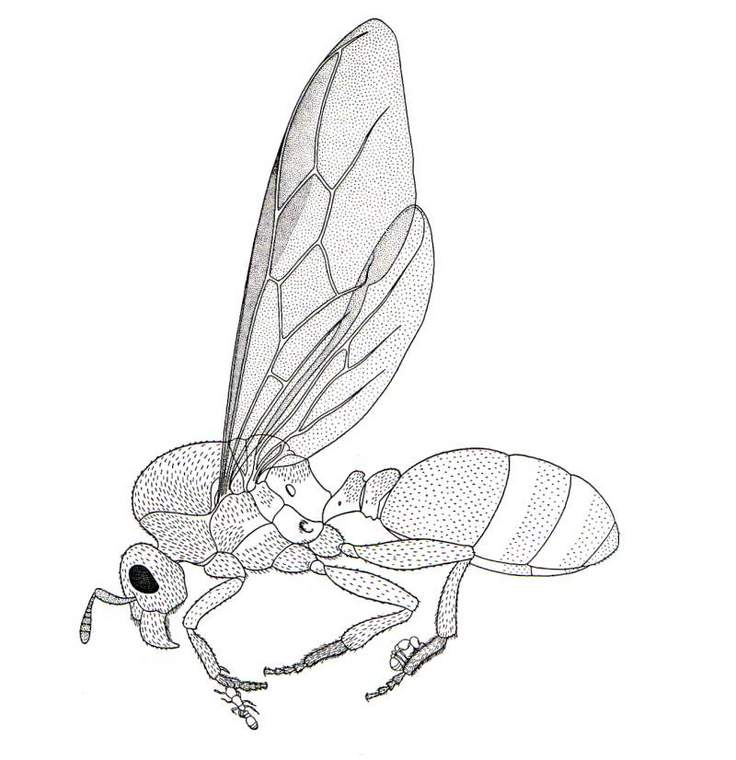 |
| Recently inseminated queen of C. vidua with (allegedly) hitchhiking sisters (Wheeler, 1936) |
 |
| Solenopsis (=Diplorhoptrum) molesta (left) with Tetramorium caespitum host (photographed by Sean McVey) |
 |
| L. limao hive on Trinidadian wall, spotted by Jo-Anne Nina Sewlal |
 |
| Lasius claviger, a temporary social parasite (Raczkowski & Luque, 2011); captured by Sharon Warner |
 |
| V. dybowskii congregated around a treat proffered by sphecologist Elmar Billig |
 |
| Newfoundlander Bombus (=Psithyrus) citrinus foraging (photographed by Mardon Erbland) |
| Portrait of slave-maker Harpagoxenus sublaevis (note prominent scrobes) |
 |
| Infanticide and revolution photographed by Alexandra Achenbach: slaves kill a slave-maker pupa |
 |
| T. tsushimae (left) battling S. koreanus (photographer unknown, due to my illiteracy in Japanese) |
Generally, the question of how, why, and from what social parasitism originated is also uncertain. Emery's Rule (named for myrmecologist Carlo Emery) in its strict sense holds that social parasites are always more closely related to their hosts than to any other taxa (Le Masne, 1956): in essence, the hypothesis holds that a given social parasite must have descended from its host (Emery, 1909). More recent data would seem to have disproved this idea—the inquiline bumblebee subgenus Psithyrus (parasites of other subgenera included in Bombus) is monophyletic, after all, as are the cuckoo paper wasps (formerly the genus Sulcopolistes; Carpenter, 1996). Cladistic analyses (Carpenter & Perera, 2006) of assorted inquilines have also poked holes in this concept's accuracy. But it remains far easier for a social insect to hoodwink its cousins and accepted into their community than it is to do the same to a more dissimilar host (Le Masne, 1970), and by and large among the Hymenoptera this holds true (social parasites are unknown so far from the Isoptera; Cervo, 2006).
 |
| Trio of Diacamma rugosum |
 |
| Queen of Anergates atratulus found in Quebec by Claude Pilon |
Both genera belong to the myrmicine tribe Tetramoriini—and both deceive the nominate genus Tetramorium: A. atratulus overwhelmingly inhabiting colonies of the familiar pavement ants in the vague T. caespitum/impurum species complex, and occasionally T. chefketi and moravicum (Lapeva-Gjonova et al., 2012); Teleutomyrmex of both species (schneideri and kutteri) are inquilines of the aforementioned pavement ant species aggregation. A. atratulus (known in the vernacular as the "dark guest ant") is widespread throughout western Eurasia, even having hitchhiked on its introduced host into North America, but remains rare throughout its range (Creighton, 1950): possibly since for whatever reason they cannot haunt queenright Tetramorium colonies (Buschinger, 2009).
 |
| Three queens of Teleutomyrmex schneideri on the back of a T. caespitum queen (Wilson, 1971) |
...Thence came the subtitle for this post, which, while contributing nothing substantive, may have grabbed your attention. Or not. Oh well.
I feel like there's some sort of holiday coming up, and that I should be wishing you well because of it...can't recall the name at the moment...
*Any organically produced chemical used for communication.
†Semiochemicals which are directed at other species and benefit the emitter only (as opposed to pheromones).
‡Literally, "stolen labor".
§With only one queen.
|| Workers that produce fertile eggs.
¶Abdomen.
____________________________________________________________
Achenbach, A. and Foitzik, S. (2009). First evidence for slave rebellion: enslaved ant workers systematically kill the brood of their social parasite Protomognathus americanus [electronic version]. Evolution, 63(4), 1068-1075. Retrieved 12/11/13 from http://onlinelibrary.wiley.com/doi/10.1111/j.1558-5646.2009.00591.x/pdf
Alloway, T. M. (1980). The origins of slavery in Leptothoracine ants (Hymenoptera: Formicidae). American Naturalist, 115, 247-261.
Apolinário, F. E. and Martius, C. (2004). Ecological role of termites (Insecta, Isoptera) in tree trunks in central Amazonian rainforests. Forest Ecology & Management, 194(1), 23-29. Retrieved 11/17/13 from http://www.aseanbiodiversity.info/Abstract/51004026.pdf
Arnold, G. (1916). A monograph of the Formicidae of South Africa. Annals of the South African Museum, 14, 1-766.
Baroni Urbani, C. (1968). Über die eigenartige Morphologie der männlichen Genitalien des Genus Diplorhoptrum Mayr (Hymenoptera: Formicidae) und die taxonomischen Schlussfolgerungen. Zoologische Morphologie Tiere, 63, 63-74.
Berg, K. (1900). Termitariophile. Comm. Mus. Nacl. Buenos Aires, 1, 212-215.
Bezerra-Gusmão, M. A.; Barbosa, J. R. C.; Barbosa, M. R. V.; Bandeira, A. G.; Sampaio, E. V. S. B. (2011). Are nests of Constrictotermes cyphergaster (Isoptera, Termitidae) important in the C cycle in the driest of semiarid caatinga in northeast Brazil? Applied Soil Ecology, 47(1), 1-5. Retrieved 11/17/13 from http://www.sciencedirect.com/science/article/pii/S0929139310002131
Blatrix, R. and Sermage, C. (2005). Role of early experience in ant enslavement: a comparative analysis of a host and a non-host species. Frontiers in Zoology, 2, 13. Retrieved 12/9/13 from http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1199612/
Breed, M. D.; McGlynn, T. P.; Stocker, E. M. and Klein, A. N. (1999). Thief workers and variation in nestmate recognition behavior in a ponerine ant, Ectatomma ruidum. Insectes Sociaux, 46(4), 327-331.
Breed, M. D.; Cook, C.; and Krasnec, M. O. (2012). Cleptobiosis in social insects. Psyche, 2012(2012), Article #484765. Retrieved 12/2/13 from http://www.hindawi.com/journals/psyche/2012/484765/#B43
Brown, W. L. (Jr.) (2000). Diversity of ants. In Agosti, D.; Majer, J. D.; and Alonso, L. E. (eds.) (280 pp.): Ants: Standard Methods for Measuring and Monitoring Biodiversity. Washington, D.C.: Smithsonian Institution Press.
Buschinger, A. (1986). Evolution of social parasitism in ants [electronic version]. Trends in Ecology and Evolution, 1, 155-160. Retrieved 12/9/13 from http://www.ncbi.nlm.nih.gov/pubmed/21227804
Buschinger, A. (1989). Evolution, speciation, and inbreeding in the parasitic ant genus Epimyrma (Hymenoptera, Formicidae) [electronic version]. Journal of Evolutionary Biology, 2, 265-283. Retrieved 12/9/13 from doi: 10.1046/j.1420-9101.1989.2040265.x.
Buschinger, A. (2009). Social parasitism among ants: a review (Hymenoptera: Formicidae) [electronic version]. Myrmecological News, 12, 219-235. Retrieved 12/24/13 from http://www.hindawi.com/journals/psyche/2012/391525/#B3
Buschinger, A.; Ehrhardt, W.; and Winter, U. (1980). The organization of slave raids in dulotic ants: a comparative study (Hymenoptera; Formicidae). Zoologie und Tierpsychologie, 53, 245-264.
Buyschinger, A.; Schlick-Steiner, B. C.; Steiner, F. M.; and Sanetra, M. (2003). Anergates atratulus, eine ungewöhnlish seltene Parasiten-Ameise. Ameisenschutz Aktuell, 17, 1-6.
Carpenter, J. M. (1996). Phylogeny and biogeography of Polistes. In Turillazzi, S. and West-Eberhard, M. J. (eds.) (pp. 18-57): Natural History and Evolution of Paper-Wasps. Oxford: Oxford University Press.
Carpenter, J. M. (1997). Phylogenetic relationships among European Polistes and the evolution of social parasitism (Hymenoptera: Vespidae, Polistinae) [electronic version]. In Grandcolas, P. (ed.): The Origin of Parasitism in Insects: Phylogenetic Tests of Evolutionary Scenarios. Mém. Mus. natn. Hist. nat., 173, 135-161. Retrieved 12/7/13 from http://research.amnh.org/iz/f/Carpenter_1997_pol.pdf
Carpenter, J. M. and Perera, E. P. (2006). Phylogenetic relationships among yellowjackets and the evolution of social parasitism (Hymenoptera: Vespidae, Vespinae) [electronic version]. American Museum Novitates, 3507, 1-19. Retrieved 12/22/13 from http://www.academia.edu/1463533/Carpenter_and_Pererra_2006_Phylogenetic_relationships_among_yellowjackets_and_the_evolution_of_social_parasitism_Hymenoptera_Vespidae_Vespinae_
Cervo, R. (2006). Polistes wasps and their social parasites: an overview [electronic version]. Annales Zoologicum Fennici, 43, 531-549. Retrieved 12/23/13 from http://www.sekj.org/PDF/anzf43/anzf43-531.pdf
Cervo, R. and Dani, F. R. (1996). Social parasitism and its evolution in Polistes. In Turillazzi, S. & Eberhard, W. M. J. (eds.) (pp. 98-112): Natural History and Evolution of Paper-Wasps. Oxford: Oxford University Press.
Cervo, R.; Lorenzi, M. C.; and Turillazzi, S. (1990). Sulcopolistes atrimandibularis, social parasite and predator of an alpine Polistes (Hymenoptera, Vespidae). Ethology, 86, 71-78.
Cervo, R.; Stemmer, C.; Castle, W.; Queller, D.; and Strassmann, J. E. (2004). Social parasitism of Polistes dominulus by Polistes nimphus (Hymenoptera, Vespidae). Insectes Sociaux, 51(2004), 101-108. Retrieved 12/4/13 from http://download.springer.com/static/pdf/949/art%253A10.1007%252Fs00040-003-0717-x.pdf?auth66=1386355114_71a8fdaa2f8883a973b3d9750d00a483&ext=.pdf
Creighton, W. S. (1950). Ants of North America. Bulletin of the Museum of Comparative Zoology, 104, 585.
Cristaldo, P. F.; Rosa, C. S.; Florencio, D. F.; Marins, A.; and DeSouza, O. (2012). Termitarium volume as a determinant of invasion by obligatory termitophiles and inquilines in the nests of Constrictotermes cyphergaster (Termitidae, Nasutitermitinae). Insectes Sociaux, 59(4), 541-548. Retrieved 11/25/13 from http://download.springer.com/static/pdf/900/art%253A10.1007%252Fs00040-012-0249-3.pdf?auth66=1385573385_dd973e696fa0d4fa0e6a92abe92c27d4&ext=.pdf
Cournault, L. and Peeters, C. (2012). Aggression regulates monogyny in non-mutilating Diacamma ants. Insectes Sociaux, 59, 533-539. Retrieved http://link.springer.com/article/10.1007%2Fs00040-012-0251-9#page-1
Couvillon, M. J.; Robinson, E, . J. H.; Atkinson, B.; Child, L.; Dent, K. R.; and Ratnieks, F. L. W. (2008). En garde: rapid shifts in honeybee, Apis mellifera, guarding behavior are triggered by onslaught of conspecific intruders. Animal Behaviour, 76(5), 1,653-1,658.
Deslippe, R. (2010). Social parasitism in ants. Social Parasitism in Ants, 3(10), 27. Retrieved 12/4/13 http://www.nature.com/scitable/knowledge/library/social-parasitism-in-ants-13256421
Dettner, K. and Liepert, C. (1996). Chemical mimicry and camouflage. Annual Review of Entomology, 39, 129-154.
D'Ettorre, P.; Errard, C.; Ibarra, F.; Francke, W.; and Hefetz, A. (2000). Sneak in or repel your enemy: Dufour's gland repellent as a strategy for successful usurpation in the slave-maker Polyergus rufescens. Chemoecology, 10, 135-142.
D'Ettorre, P. and Heinze, J. (2001). Sociobiology of slave-making ants [electronic version]. Acta Ecologica, 3, 67-82. Retrieved 12/9/13 from http://download.springer.com/static/pdf/995/art%253A10.1007%252Fs102110100038.pdf?auth66=1386814388_8c7addffa306c41c5627eb73e1f4b13f&ext=.pdf
Emery, C. (1909). Über den ursprung der dulotischen, parasitischen und myrmekophilen Ameisen. Biol. Centralbl., 29, 352-362.
Espadaler, X.; Gómez, C.; and Suñer, D. (1995). Seed-robbing between ant species intervenes in the myrmecochory of Euphorbia characias (Euphorbiaceae). Psyche, 102(1-2), 19-25.
Ettershank, G. (1966). A generic revision of the world Myrmicinae related to Solenopsis and Pheidologeton (Hymenoptera: Formicidae). Australian Journal of Zoology, 14, 73-171.
Fisher, R. M. (1983). Behavioural interactions between a social parasite, Psithyrus citrinus (Hymenoptera: Apidae) and its bumble bee hosts. Proceedings of the Entomological Society of Ontario, 114, 55-60.
Fisher, R. M. and Sampson, B. J. (1992). Morphological specializations of the bumble bee social parasite Psithyrus ashtoni (Cresson) (Hymenoptera: Apidae). Canadian Entomology, 124, 69-77.
Florencio, D. F.; Marins, A.; Rosa, C. S.; Cristaldo, P. F.; Araújo, A. P. A.; Silva, I. R.; and DeSouza, O. (2013). Diet Segregation between Cohabiting Builder and Inquiline Termite Species. PLoS One, 8(6), e66535. Retrieved 11/29/13 from http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0066535#pone.0066535-Schnrogge1
Forel, A. (1898). La parabiose chez les fourmis. Bull. Soc. Vaud. Sci. Nat. , 34, 380-384.
Gösswald, K. (1953). Histologische Untersuchungen an der arbeiterlosen Ameise Teleutomyrmex schneideri Kutter (Hym. Formicidae). Mitteilungen der Schweizerischen Entomologischen Gesellschaften, 26(2), 81-128.
Grasso, D. A.; Mori, A.; Giovannotti, M.; and Le Moli, F. (2004). Interspecific interference behaviors by workers of the harvesting ant Messor capitatus (Hymenoptera Formicidae). Ethology, Ecology & Evolution; 16(3), 197-207.
Heinze, J.; Lautenschläger, B.; and Buschinger, A. (2007). Female-biased sex ratios and unusually potent males in the social parasite Anergates atratulus (Hymenoptera: Formicidae). Myrmecological News, 10, 1-5. Retrieved 12/24/13 from http://www.myrmecologicalnews.org/cms/images/pdf/volume10/mn10_1-5_non-printable.pdf
Hölldobler, B. (1986). Food robbing in ants, a form of interference competition. Oecologia, 69(1), 12-15.
Hölldobler, B. and Wilson, E. O. (1990). The Ants. Cambridge: the Belknap University Press of Harvard University Press.
Huber, P. (1810). Recherches sur les Moeurs ded Fourmis Indigènes. Paris: J. J. Paschoud.
Jeral, J. M.; Breed, M. D.; and Hibbard, B. E. (1997). Thief ants have reduced quantities of cuticular compounds in a ponerine ant, Ectatomma ruidum. Physiological Entomology, 22(3), 207-211.
Lapeva-Gjonova, A.; Kiran, K.; and Aksoy, V. (2012). Unusual ant hosts of the socially parasitic ant Anergates atratulus (Schenck, 1852) (Hymenoptera, Formicidae). Psyche, 2012(2012), 1-3. Retrieved 12/24/13 from http://www.hindawi.com/journals/psyche/2012/391525/#B4
Le Masne, G. (1956). Recherches sur les fourmis parasites: Plagiolepis grassei et l'evolution des Plagiolepis parasites. C. R. Acad. Sci., 243, 673-675.
Le Masne, G. (1970).
Le Moli, F. (1980). On the origin of slaves in dulotic ant societies. Boll. Zool., 47, 207-212.
Lenoir, A.; D'Ettorre, P.; Errard, C.; and Hefetz, A. (2001). Chemical ecology and social parasitism in ants. Annual Review of Entomology, 46, 573-599.
Lepage, M. G. and Darlington, J. P. E. C. (1984). Observations on the ant Carebara vidua F. Smith preying on termites in Kenya [electronic version]. Journal of Natural History, 18(2), 293-302. Retrieved 11/29/13 from http://www.ingentaconnect.com/content/tandf/tnah/1984/00000018/00000002/art00008
Longino, J. T. (2004). Carebara of Costa Rica. Retrieved 11/30/13 from http://academic.evergreen.edu/projects/ants/genera/carebara/home.html
Lorenzi, M. C.; Caldi, M.; and Cervo, R. (2007). The chemical strategies used by Polistes nimphus social wasp usurpers (Hymenoptera: Vespidae) [electronic version]. Biological Journal of the Linnaean Society, 91, 505-512. Retrieved 12/6/13 from http://eligreenbaum.iss.utep.edu/Wasp%20Parasitism.pdf
Lowe, G. H. (1948). Some observations on the habits of a Malayan ant of the genus Carebara. Proceedings of the Royal Entomological Society of London (A), 23, 51-53.
MacDonald, J. F. (1989). An exposed Vespula germanica colony usurped by maculifrons. Sphecos, 18, 9-10.
Maschwitz, U.; Dorow, W. H. O.; Buschinger, A.; and Kalytta, G. (2000).
Maschwitz, U.; Liefke, C.; and Buschinger, A. (2001).
Maschwitz, U.; Go, C.; Kaufmann, E.; and Buschinger, A. (2004).
Mathews, A. G. A. (1977). Studies on Termites from the Mato Grosso State, Brazil. Rio de Janeiro: Academia Brasilieria de Ciências.
Matsura, M. and Yamane, S. (1990). Biology of the Vespine Wasps. Berlin: Springer Verlag.
Michener, D. C. (2000). The Bees of the World. Baltimore: Johns Hopkins University Press.
Pamminger, T.; Leingärtner, Achenbach, A.; Kleberg, I.; Pennings, P. S.; and Foitzik, S. (2013). Geographic distribution of the anti-parasite trait "slave rebellion". Evolutionary Ecology, 27, 39-49. Retrieved 12/11/13 from http://download.springer.com/static/pdf/933/art%253A10.1007%252Fs10682-012-9584-0.pdf?auth66=1386977538_7c04b8eb10d5042f72d42029e02505b6&ext=.pdf
Perfecto, I. and Vandermeer, J. H. (1993). Cleptobiosis in the ant Ectatomma ruidum in Nicaragua. Insectes Sociaux, 40(3), 295-299.
Raczkowski, J. M. and Luque, G. M. (2011). Colony founding and social parasitism in Lasius (Acanthomyops). Insectes Sociaux, 58, 237-244.
Regnier, F. E. and Wilson, E. O. (1971). Chemical communication and "propaganda" in slave-maker ants. Science, 172, 267-269.
Robertson, H. G. and Villet, M. H. (1989). Colony founding in the ant Carebara vidua: the dispelling of a myth [electronic version]. Suid-Afrikaanse Tydskrif vir Wetenskap, 85, 121-123. Retrieved 11/30/13 from https://archive.org/stream/ants_04173/4173#page/n1/mode/2up
Sakagami, S. F. and Fukushima, K. (1957). Vespa dybowskii André as a facultative temporary social parasitism [electronic version]. Insectes Sociaux, 4(1), 1-12. Retrieved 12/7/13 from http://link.springer.com/article/10.1007%2FBF02226244
Sakagami, S. F. and Laroca, S. (1963). Additional observations on the habits of the cleptobiotic stingless bee, the genus Lestrimelitta Friese (Hymenoptera, Apoidea). Journal of the Faculty of Science, 15(2), 319-339.
Sakagami, S. F.; Roubik, D. W.; and Zucchi, R. (1993). Ethology of the robber stingless bee, Lestrimelitta limao (Hymenoptera, Apidae). Sociobiology, 21(2), 237-277.
Sanetra, M. and Güsten, R. The socially parasitic genus Strongylognathus Mayr in North Africa (Insecta: Hymenoptera: Formicidae). Zootaxa, 20, 1-20.
Stuart, R. J. and Alloway, T. M. (1983). The slave-making ant, Harpagoxenus canadensis M. R. Smith, and its host species, Leptothorax muscorum (Nylander): slave raiding and territoriality. Behaviour, 85, 58-90.
Stumper, R. (1950). Les associations complexes des fourmis: commensalisme, symbiose et parasitisme. Bulletin Biologique de la France et de la Belgique, 84(4), 376-399.
Taylor, L. H. (1939). Observations on social parasitism in the genus Vespula Thomson. Annals of the Entomological Society of America, 32, 304-315.
Thompson, C. R. (1980). Monograph of the Solenopsis (Diplorhoptrum) of Florida. Ph.D. dissertation. University of Florida.
Thompson, C. R. (1989). The thief ants, Solenopsis molesta group, of Florida (Hymenoptera: Formicidae). Florida Entomologist, 72(2), 268-291. Retrieved 11/30/13 from http://journals.fcla.edu/flaent/article/view/58486/56165
Turillazzi, S. (1991). The Stenogastrinae. In Ross, K. G. and Matthews, R. W. (eds.) (pp. 74-98): The Social Biology of Wasps. Ithaca: Cornell University Press.
Topoff, H. and Zimmerli, E. (1991). Formica wheeleri: Darwin's predatory slave-making ant [electronic version]? Psyche, 98(4), 309-317. Retrieved 12/11/13 from http://www.hindawi.com/journals/psyche/1991/034829/abs/
Vasconcellos, A.; Araújo, V. F. P.; Moura, F. M. S.; and Bandeira, A. G. (2007). Biomass and population structure of Constrictotermes cyphogaster (Silvestri) (Isoptera: Termitidae) in the dry forest of Caatinga, Northeastern Brazil. Neotropical Entomology, 36, 693-698.
Wesson, L. G. (1939). Contributions to the natural history of Harpagoxenus americanus Emery (Hymenoptera; Formicidae). Transactions of the American Entomological Society, 65, 97-122.
Wheeler, W. M. (1901). The compound and mixed nests of American ants. Part II. The known cases of social symbiosis among American ants. American Naturalist, 35, 513-539.
Wild, A. (October 24, 2013). Polyergus moves forward, and a modest proposal for kidnapper ants. Retrieved 12/24/13 from http://www.myrmecos.net/2013/10/24/polyergus-moves-forward-and-a-modest-proposal-for-kidnapper-ants/
Wilson, E. O. (1971). The Insect Societies. Cambridge: Harvard University Press.
Wilson, E. O. (1975). Leptothorax duloticus and the beginnings of slavery in ants [electronic version]. Evolution, 29(1), 108-119. Retrieved 12/11/13 from http://gap.entclub.org/taxonomists/Wilson/Leptothorax%20duloticus.pdf
Wilson, E. O. (2003). Pheidole in the New World: a Dominant, Hyperdiverse Ant Genus. Cambridge: Harvard University Press.
Witte, V.; Lehmann, L.; Lustig, A.; and Maschwitz, U. (2009). Polyrhachis lama, a parasitic ant with an exceptional mode of social integration. Insectes Sociaux, 56(3), 301-307. Retrieved 12/23/13 from http://link.springer.com/article/10.1007/s00040-009-0024-2
Yamaguchi, T. (1995). Intraspecific competition through food robbing in the harvester ant, Messor aciculatus (Fr. Smith),and its consequences on colony survival. Insectes Sociaux, 42(1), 89-101.
Zimma, B. O. (2002). Identification of a repellent allomone in the social parasitic bumblebee Psithyrus norvegicus (Hymenoptera, Apidae). Diploma thesis, University of Vienna.
Zimma, B. O.; Ayasse, M.; Tengö, J.; Ibarra, F.; Schulz, C.; Francke, W. (2003). Do social parasitic bumblebees use chemical weapons? (Hymenoptera, Apidae). Journal of Comparative Physiology, A; 189(10), 769-775. Retrieved 12/8/13 from http://download.springer.com/static/pdf/779/art%253A10.1007%252Fs00359-003-0451-x.pdf?auth66=1386699090_2c33d50d4dc2d2f9905fda2b63d1f4da&ext=.pdf
No comments:
Post a Comment